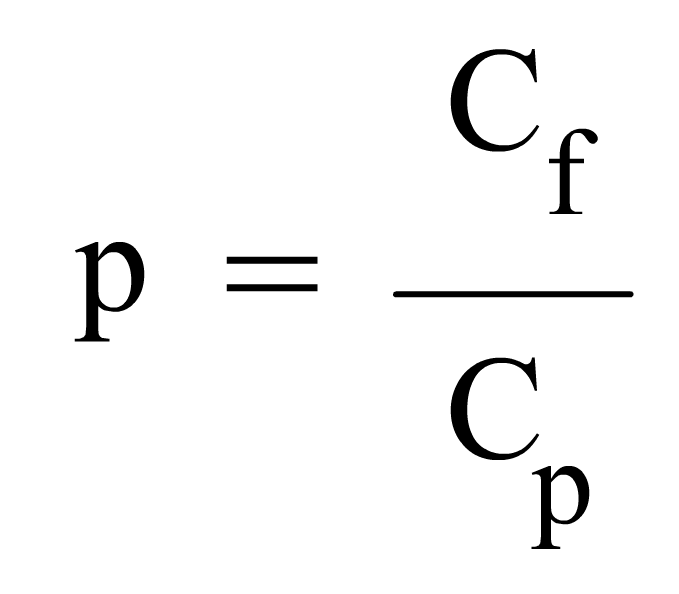
|
P
|
ARENTESCO Y CONSANGUINIDAD (2) |
|
www.labordadurtx.org
>> artículos >> parentesco (2)
|
En el artículo anterior definimos algunos de los conceptos básicos para poder comprender ahora mejor el significado de parentesco y consanguinidad. No obstante y teniendo en cuenta que el estudio de la transmisión de los genes se basa en el cálculo de probabilidades, no está de más que repasemos sus distintas definiciones.
| PROBABILIDAD SIMPLE: |
Para definir este concepto podemos razonar de la siguiente manera:
Cada vez que arrojamos al aire una moneda, lo mismo puede salir cara que cruz, y como esto sucede todas las veces, decimos que es igualmente probable que salga cara que cruz o sea que la probabilidad es la mísma. Entonces como que de dos casos posibles, solamente se ha verificar uno en cada tirada, decimos que la probabilidad de que salga cara o cruz es 1/2.
| Pongamos otro ejemplo: Si suponemos el caso del dado, al arrojarlo al aire se dan seis posibilidades, todas ellas equiprobables, a saber: que salga el 1, el 2, el 3, el 4 el 5 o el 6. Luego la probabilidad de cada una será 1/6. |
En general, la probabilidad de un suceso que se presenta en "C" casos favorables sobre "C", casos posibles, se hallará por el cociente:
|
|
si un suceso se presenta siempre, su probabilidad es uno y decimos de él que es seguro; si no se presenta nunca, su probabilidad es cero y decimos de él que es imposible.
En el caso de la transmisión genética, cada individuo traspasa siempre sólo la mitad de sus genes o sea, uno de cada dos cromosomas homólogos, y como que ello se produce al azar, la probabilidad de que traspase unos u otros será 1/2.
| PROBABILIDAD COMPUESTA: |
Es la probabilidad de un acontecimiento constituido por una sucesión de otros acontecimientos probables que se desarrollan independientemente unos de otros pero que estan relacionados entre sí. La probabilidad resultante final es igual al producto de las probabilidades de cada uno.
| Por ejemplo, en la transmisión de los genes la probabilidad de que un individuo reciba unos genes determinados de un antepasado suyo, viene dada por el producto de las probabilidades de transmisión de estos genes en cada generación, desde el antepasado hasta este individuo. |
Vemos que el paso de los genes de una generación a otra es un hecho independiente en cada generación pero el conjunto de genes del individuo resultante depende de como se han transmitido los genes desde el antepasado hasta llegar a él.
Como que la probabilidad de transmisión de los genes en cada generación siempre es 1/2, bastará multiplicar esta cifra por si misma, tantas veces como generaciones separen a este individuo de su antepasado.
Una vez hecho este pequeño repaso del cálculo de probabilidades ya podemos desarrollar los conceptos de consanguinidad y parentescos.
| CONSANGUINIDAD: |
La consanguinidad se puede definir como el cruzamiento entre individuos emparentados. Cuando mayor sea el parentesco entre dos individuos, mayor será el coeficiente de consanguinidad de su progenie y menor será la probabilidad de que esta progenie sea heterocigótica
| COEFICIENTE DE CONSANGUINIDAD: |
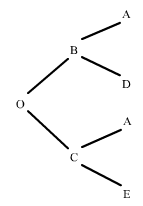
Comenzaremos considerando la situación más sencilla posible: dos descendientes B y C que tienen un progenitor A en común. Utilizando flechas para indicar la dirección de la herencia, esta situación se puede representar de la siguiente forma:
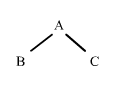 |
|
figura 1
|
Se dice que A es el antepasado común de B y C. Si B y C reciben ambos una copia del mismo gen, es decir, el mismo segmento de ADN de un locus cualquiera de A, podemos decir que B y C tienen genes que son idénticos por descendencia procedentes de A. Recordemos que dos genes son identicos por descendencia si son copias del mismo segmento de ADN.
Supongamos ahora que B y C se cruzan para producir un descendiente O:
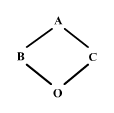 |
|
figura 2
|
Si B y C tienen genes que son idénticos por descendencia, en cualquier locus existe una cierta probabilidad de que O herede dos genes que son idénticos por descendencia, uno procedente de A a través de B y otro procedente de A a través de C. Ahora estamos ya en disposisición de definir el coeficiente de consanguinidad:
"El coeficiente de consanguinidad de un individuo es la probabilidad de que dos genes presentes en un locus de dicho individuo sean idénticos por descendencia".
El coeficiente de consanguinidad se designa generalmente mediante la letra F. Sin un individuo tiene dos genes que son idénticos por descendencia en un determinado locus, será homocigoto para dicho locus. Por tanto, el coeficiente de consanguinidad es un reflejo de la homocigosis.
| LA POBLACION BASE. |
En genética se define como población a un grupo de individuos que se cruzan entre sí y comparten un acervo común de genes que se transmiten de generación en generación de acuerdo con las leyes de Mendel. Para medir el coeficiente de consanguinidad se toma como base a un grupo de individuos, -de ahí el nombre de población base-, que se supone no estan emparentados.
Es importante tener en cuenta que el coeficiente de consanguinidad no mide la homocigosis en un sentido absoluto. De hecho, es una cantidad relativa que lo que realmente mide es el descenso de la heterocigosis en relación a una población base en la que todos los individuos se supone que no estan relacionados y que tienen una consanguinidad cero. En otras palabras mide el grado en que un individuo es menos heterocigoto que los individuos que se supone tienen consanguinidad cero. En la práctica, suponemos que tienen consanguinidad cero (F=0), todos aquellos individuos sobre los que no disponemos de información.
Para calcular el descenso de la heterocigosis se resta de la unidad (heterocigosis total) el coeficiente de consanguinidad F: (1-F).
Sin embargo, no olvidemos que por el sólo hecho de que el Montaña es raza pura, los individuos que conformen la población base de la cual partamos en cada caso, ya llevan implicito un elevado nivel de homocigosis en muchos de sus loci, producto de una consanguinidad ancestral, pero ello no importa; los homocigotos en esta población base tienen genes que son considerados como "similares en estado" en vez de idénticos por descendencia. Esto es, son indénticos en estructura porque tienen la misma secuencia de ADN, y aún así, no son idénticos por descendencia porque proceden de diferentes genitores. Dicho de otro modo, son homocigotos para copias de distintos genes que proceden de genitores ancestrales, en vez de para dos copias de un único gen originario de un genitor ancestral.
Entonces, es lícito que nos preguntemos como las medidas de parentesco y consanguinidad pueden ser de alguna utilidad cuando su magnitud real depende de la elección arbitraria de una población base, cuyos miembros se supone por conveniencia que no tienen antepasados en común.
A primera vista, es difícil reconciliar esto con el hecho de que si retrocedemos un suficiente número de generaciones, dos individuos cualesquiera deben tener antepasados en común y ser por tanto parientes. En la práctica, esto no representa un problema siempre y cuando recordemos que los coeficientes de parentesco y consanguinidad son cantidades relativas en vez de absolutas. Así, si un individuo tiene un coeficiente de consanguinidad cero relativo a una determinada población base, la probabilidad de que este individuo sea heterocigoto en un locus cualquiera es igual a la frecuencia de heterocigotos en dicho locus en la población base. Si el nivel de heterocigosis en la población base es bastante bajo, debido por ejemplo a un número significativo de cruzamientos entre parientes próximos entre los antepasados de la población base, el individuo con un coeficiente de consanguinidad cero tendrá de hecho un nivel bastante bajo de heterocigosis. El punto importante es, sin embargo, que si un segundo individuo tiene un coeficiente de consanguinidad de 0,3 en relación a la misma población base, inmediatamente sabemos que este segundo individuo es un 30% menos heterocigoto que el individuo cuyo coeficiente de consanguinidad era cero.
Por otro lado, sin un individuo tiene un coeficiente de consanguinidad de F=1, la probabilidad de que este individuo sea heterocigoto en un locus cualquiera, es igual a cero (1-1=0); puesto que el individuo tienen genes idénticos por descendencia en todos los loci, ha de ser necesariamente homocigoto en todos ellos.
Esto subraya el hecho de que los coeficientes de consanguinidad permiten comparar los niveles relativos de heterocigosis entre individuos cuyos pedigríes pueden seguirse hacia atrás hasta la misma población base. Más adelante desarrollaremos la fórmula para calcular el coeficiente de consanguinidad.
| PARENTESCO DIRECTO: |
El parentesco más simple es aquel que hay entre un progenitor (P) y su descendiente (N). En un locus cualquiera un descendiente tiene dos genes, uno de los cuales (el gen paterno) es una copia de uno de los genes de su padre, y el otro (el gen materno) es una copia de uno de los genes de su madre. Puesto que esto es cierto para todos los loci, podemos decir que un descendiente comparte exactamente 1/2 de sus genes con cada uno de sus padres. En otras palabras, un progenitor y un descendiente tienen exactamente 1/2 de sus genes en común.
Luego podemos definir que: "La proporción esperada de genes en común entre dos individuos no consanguíneos es el parentesco entre ellos". (Si lo individuos son consanguíneos hemos de aplicar la definición que veremos más adelante en el parentesco colateral). Por tanto el parentesco entre un progenitor y su descendiente es 1/2, (probabilidad simple). Tanto el parentesco directo como el parentesco colateral, que estudiaremos más adelante, se designan con la letra "a".
Ahora, hagámonos la siguiente pregunta: ¿Cuál es el parentesco entre un descendiente (O) y su abuelo (A). Para contestarla debemos de seguir la transmisión de los genes desde el abuelo (A) al padre (P) y al descendiente (O). Es evidente que (A) transmite exactamente 1/2 de sus genes a (P), que a su vez transmite exactamente 1/2 de sus genes a (O). De aquí que como promedio (O) recibirá 1/2 x 1/2 = 1/4 de sus genes de (A) (probabilidad compuesta). Por tanto el parentesco entre un descendiente y uno cualquiera de sus abuelos es 1/4.
No obstante, es importante advertir que mientras que la proporción real de genes en común entre un descendiente y su progenitor es siempre exactamente 1/2, la proporción real de genes en común entre un descendiente y su abuelo, y entre todos los restantes parientes, puede no ser exactamente igual a la proporción esperada.
Por ejemplo, la proporción real de genes en común entre un determinado descendiente y uno de sus cuatro abuelos podría ser mayor o menor que 1/4. Esta proporción podría ser 0 (si se diese el caso de que el gameto procedente de (P) no hubiese llevado ninguno de los genes que (P) heredó de (A) o podría alcanzar el valor de 1/2 (si por azar el gameto procedente de (P) hubiese llevado todos los genes que (P) heredó de (A)).
No hay forma de saber la proporción real de genes en común entre un determinado descendiente y uno de sus cuatro abuelos. Lo único que podemos decir es que la proporción esperada, es decir el parentesco, es 1/4.
Mediante un razonamiento semejante al que acabamos de utilizar, se puede deducir que el parentesco entre un descendiente y uno de sus bisabuelos es:
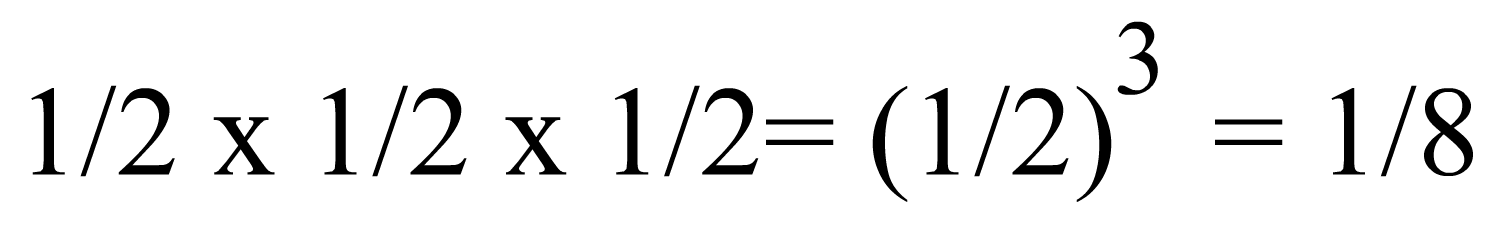 |
En general, el parentesco entre un individuo y uno de sus antepasados disminuye en 1/2 por cada generación que lo separa de dicho antepasado.
Por tanto, el parentesco entre un individuo y un antepasado cualquiera es:
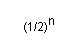 |
donde "n" es el número de generaciones entre el individuo y su antepasado en la línea de descendencia directa del antepasado al individuo.
Se deduce que independientemente de lo sobresaliente que pueda haber sido un determinado antepasado, si sólo aparece una vez y varias generaciones atrás en el árbol genealógico de un individuo, existe sólo una pequeña probabilidad de que el individuo en cuestión (al que llamamos O) haya heredado alguno de los genes de su antepasado. De aquí que sea inutil seleccionar un individuo solamente porque su pedigrí contiene un antepasado distante que se da la circunstancia de que fué muy famoso.
La situación es ligeramente distinta si un determinado antepasado aparece más de una vez en el árbol genealógico. En este caso, cada aparición del antepasado en el pedigrí proporciona una oportunidad independiente de que el individuo (O) haya heredado los genes del antepasado. El parentesco total es, en este caso, igual a la suma de las contribuciones independientes hechas por cada línea de descendencia directa.
Por ejemplo:
 |
|
figura 3
|
Observemos que el individuo B es padre y a la vez abuelo de O. ¿Cual será el parentesco total entre ellos?. En este caso hay dos líneas de descendencia directas, por tanto, tal como hemos enunciado, sus contribuciones al parentesco serán la suma de ambas:
 |
El tipo de parentesco que hemos considerado se denomina "parentesco directo" ya que se refiere al parentesco que puede seguirse en una línea directa o línea de descendencia de antepasado a descendiente. El otro tipo de parentesco es el que existe entre dos individuos que tienen un antepasado en común y pueden por tanto haber heredado el mismo gen de dicho antepasado. Este tipo de parentesco se denomina "parentesco colateral".
| PARENTESCO COLATERAL: |
Consideremos dos descendientes, (B) y (C), con un progenitor (A) en común, (figura 1). En este caso, (B) y (C) se dice que son "medio hermanos" y (A) es su antepasado común. Si (B) y (C) reciben ambos una copia del mismo gen de un locus cualquiera de (A), podemos decir que (B) y (C) tienen genes que son idénticos por descendencia procedentes de (A).
Ahora podemos considerar una definición más general del parentesco: "El parentesco entre dos individuos cualesquiera es el número esperado de genes en un locus de un individuo que son idénticos por descendencia con un gen elegido al azar en el mismo locus del otro individuo".
Esta definición es más correcta que la que hemos usado en el parentesco directo, tanto si los individuos emparentados son consanguíneos como si no. Si los individuos emparentados no son consanguíneos, esta definición es equivalente a la que se ofreció en el parentesco directo. Para comprender mejor la diferencia entre las dos definiciones observemos la siguienta tabla:
|
Tipo de Gen
|
Coeficiente de Consanguinidad
|
Parentesco aditivo entre B yC
|
|||
|
B
|
C
|
FB
|
FC
|
||
|
Definición exacta
|
Definición simple
|
||||
|
A1A2
|
A3A4
|
0
|
0
|
0
|
0
|
|
A1A2
|
A3A2
|
0
|
0
|
1/2
|
1/2
|
|
A1A2
|
A1A2
|
0
|
0
|
1
|
1
|
|
A1A1
|
A1A1
|
1
|
1
|
1
|
2
|
|
tabla 1
|
|||||
|
A1
|
A2
|
|
|
A1
|
A1A1
|
A1A2
|
|
A2
|
A2A1
|
A2A2
|
|
tabla 2
|
Vemos que la probabilidadd de que un descendiente tenga dos copias del mismo gen es 1/4 para A1A1 y 1/4 para A2A2, lo que da lugar a un coeficiente de consanguinidad F = 1/4 + 1/4 = 1/2.
Como en el caso de los parientes directos, el parentesco entre dos parientes colaterales disminuye 1/2 por cada generación que separa los dos parientes a lo largo de un determinado trayecto. La única diferencia es que en el caso de los parientes colaterales, el trayecto entre ellos consta de dos líneas de descendencia: una desde el antepasado común hasta uno de los parientes y la otra desde el antepasado común hasta el otro pariente. Para subrayar esto, el número total de generaciones que separa a los dos parientes colaterales se escribe n + n', dónde n es el número de generaciones entre el antepasado común y el primer pariente y n' es el número de generaciones entre el antepasado común y el segundo pariente. Así para un trayecto cualquiera, el parentesco entre dos parientes colaterales es:
A veces, puede haber más de un trayecto que pasa a través de un antepasado común, y a veces puede haber más de un antepasado común. En todos los casos, el parentesco total es la suma del parentesco con el que contribuye cada uno de los trayectos.
El antepasado común tiene evidentemente una importancia crítica en la determinación de los parentescos colaterales. ¿Qué ocurre cuando este antepasado es consanguíneo?. Hemos visto que el coeficiente de consanguinidad de un individuo es la probabilidad de que dos genes en un locus de dicho individuo sean indénticos por descendencia. Se deduce que cuanto más alto sea el coeficiente de consanguinidad del antepasado común, mayor será la probabilidad de que transmita el mismo gen a cada uno de los dos parientes colaterales que descienden de él, y por tanto mayor será la probabilidad de que los dos parientes tengan ambos genes que son idénticos por descendencia. De hecho, si el antepasado común es consanguíneo, el parentesco entre los dos parientes colaterales que descienden de él, aumenta en una proporción igual al coeficiente de consanguinidad del antepasado común. Esto significa que el trayecto del antepasado común consanguíneo deberá contener un término añadido en consideración a su endogamia. Algebraicamente, esto equivale a:
|
trayecto + (trayecto x FA) = trayecto (1+FA)
|
lo que representa multiplicar (1/2)n+n' para cada trayecto del antepasado común consanguíneo por (1+FA), donde FA es el coeficiente de consanguinidad del antepasado común. Teniendo todo esto en cuenta, y utilizando el símbolo a para el parentesco, podemos expresar ahora el parentesco entre dos individuos cualesquiera como:
donde el sumatorio (letra sigma mayúscula del alfabeto griego) significa que ha de efectuarse la suma de los distintos trayectos (p), y en caso de que el antepasado fuera consanguíneo, antes de incluir su trayecto en la suma, se ha de multiplicar por (1+Fa).
La misma fórmula puede usarse también para calcular el parentesco directo, simplemente teniendo en cuenta que en tales casos sólo existe una línea de descendencia para cada trayecto, y por tanto n'=0 para cada trayecto.
| CALCULO DEL COEFICIENTE DE CONSANGUINIDAD: |
Sigamos con el ejemplo de la figura 2, con el de cruzamiento entre B y C para producir un descendiente O y consideremos un gen elegido al azar que B transmite a O.
Para calcular el coefiente de consanguinidad de O, necesitamos calcular la probabilidad de que el otro gen del mismo locus en O (el gen procedente de C) sea idéntico por descendencia al gen recibido de B.
Para calcular esta probabilidad hemos de recordar, en este caso, que el parentesco entre B y C es el número esperado de genes de C que son idénticos por descendencia con un gen elegido al azar en el mismo locus de B.
Como que C es el otro progenitor de O y por tanto existe una probabilidad de 1/2 de que un gen cualquiera de C sea transmitido a O, se deduce que la probabilidad de que O reciba un gen de C que sea idéntico por descendencia con el gen que O recibió de B, es igual al parentesco entre B y C, multiplicado por 1/2.
Esto significa que: "El coeficiente de consanguinidad de un individuo cualquiera es 1/2 del parentesco entre los padres de este individuo".
Todo esto puede parecer muy complicado pero con unos ejemplos lo veremos mucho más claro:
 |
|
figura 4
|
Es mucho más fácil si modificamos el árbol genealógico tradicional para convertirlo en un diagrama de trayectos, en el que los individuos implicados aparecen solamente una vez y en el que las flechas indican la dirección de la herencia (fig. 2):
|
|
figura 5
|
Obsérvese que en el diagrama no aparecen todos los individuos; se han excluido aquellos que no son antepasados comunes y que no se encuentran en ningún trayecto entre el antepasado común y el individuo en cuestión (en este caso D y E).
A continuación calcularemos el parentesto entre los dos medios hermanos, B y C, y el coeficiente de consanguinidad de su descendiente O. Comenzaremos por construir la siguiente tabla:
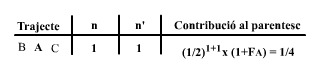 |
En esta tabla, la A se ha subrayado para indicar que se trata del antepasado común en este trayecto. Como en el diagrama hay un solo trayecto, existe una única entrada (fila) de datos en la tabla que nos proporciona el parentesto entre B y C como:
|
figura 6
|
Se deduce por tanto que el coeficiente de consanguinidad de O viene dado por:
|
figura 7
|
Hasta aquí se han expuesto las definiciones y las fórmulas matemáticas para llegar a una comprensión razonada lo más clara posible de los conceptos tratados. No obstante, una vez comprendidas estas nociones ya estamos en disposición de aplicar un metódo abreviado para llegar a los mismos resultados. Para ello podemos resumir en una, las fórmulas de las figuras 6 y 7:
|
figura 8
|
Siendo la suma de todos los trayectos posibles y n, el número de generaciones transcurridas del antepasado común desde el lado paterno al lado materno, y FA es el coeficiente de endogamia del antepasado común A. A es un símbolo general representativo de cualquiera de los antepasados susceptibles de contribuir a la endogamia de X. Si los antepasados comunes no son endógamos, entonces FA = 0 y la fórmula se escribe:
|
|
|
figura 9
|
Veamos ahora otro ejemplo en un pedigrí algo más complejo:
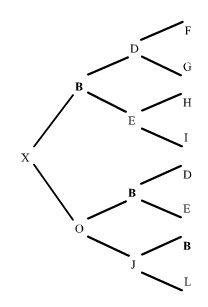 |
|
figura 10
|
El individuo X es el resultante de dos cruzamientos consecutivos respecto al padre B. En la ascendencia la endogamia se produce en dos momentos. En el primero, el apareamiento de J con B que da el trayecto XOJ; en el segundo, el apareamiento de O con B que dá el trayecto XO.
Ahora procedamos de la siguiente manera:
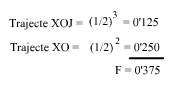 |
¿Que hemos hecho? En el primer trayecto hemos contado el número de generaciones transcurridas de B a X por el lado paterno, y vuelta a B por el lado materno hasta encontrar el primer cruce de B, dándonos tres generaciones, o sea n=3, (reparemos que el antepasado común B nunca se cuenta).
En el segundo trayecto hemos obrado de igual manera dándonos dos generaciones, o sea n=2. Luego, solamente nos ha faltado sumar estas endogamias parciales.
Observemos que tanto D como E, aún estando a ambos lados del pedigrí (paterno y materno) no se cuentan, puesto que sus contribuciones a la endogamia están incluidas en la de B. Observemos también, que este coeficiente de consanguinidad F=0.375 del individuo X, es justamente la mitad del parentesco que hay entre sus padres (a=0,75), Fig. 3.
Esto viene a corroborar lo dicho cuando hemos explicado el cálculo del coeficiente de consanguinidad: "El coeficiente de consanguinidad de un individuo es siempre 1/2 del parentesco entre sus padres".
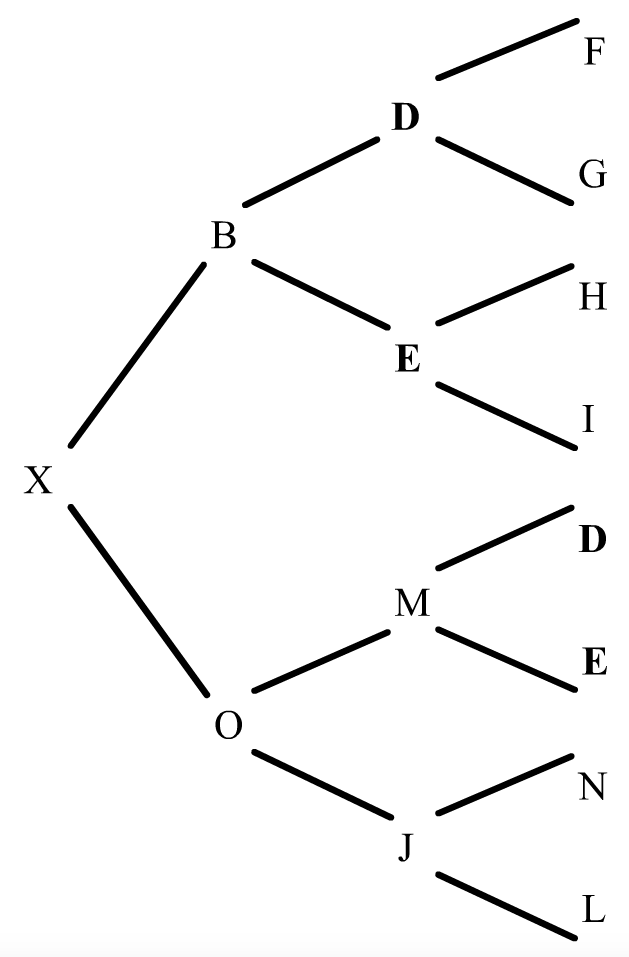
Qué ocurre si en el pedigrí de la figura 10 se sustituye la segunda aparición de B por un hermano M, y la tercera por un individuo N sin ningún parentesco respecto a B?. Esto significa que B no contribuye a la endogamia de X, en cambio D y E sí lo hacen. Veamos el ejemplo:
 |
|
figura 11
|
El trayecto para D es BXOM = (1/2)4 = 0.0625 y el de E también es BXOM = (1/2)4 = 0.0625. Intervienen dos antepasados diferentes y la endogamia total (Fx) es la suma de los dos trayectos; a saber, 0,125. Este ejemplo demuestra que puede haber más de un antepasado común en un pedigrí y que cada uno de ellos hará su propia aportación al coeficiente de endogamia (Fx).
Una vez vistos estos ejemplos, podemos resumir en tres reglas básicas la determinación de los caminos (trayectos) genealógicos:
1ª. El antepasado común del individuo X nunca se cuenta.
2ª. Identificar siempre todos los caminos (trayectos) posibles.
3ª. Los individuos presentes a ambos lados del pedigrí, nunca serán contados más de una vez en el trayecto.
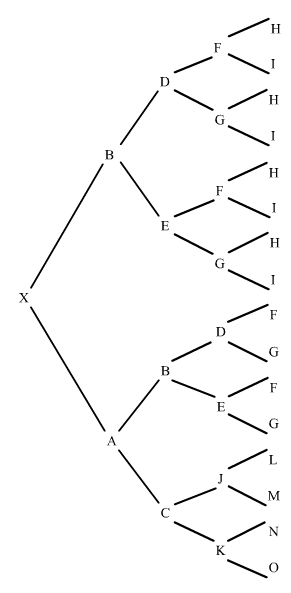
Ahora vamos a calcular el coeficiente de consanguinidad de un individuo X, en el que el ascendiente común B es endógamo (hijo de hermanos, siendo sus abuelos también hermanos) y es a la vez, padre y abuelo del individuo X:
 |
|
figura 12
|
El trayecto de B es XA o sea (1/2)2, pero al ser B endógamo hemos de calcular su coeficiente de consanguinidad aparte:
 |
aplicando la fórmula:
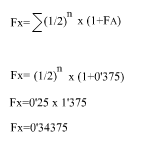 |
Luego, el índice de consanguinidad de X es:
|
Fx= 0'34375 o el que és el mateix 34'375%.
|
Otro ejemplo complicándolo un poquito más. Vamos a sustituir al individuo C del ejemplo anterior por E que es la madre de B:
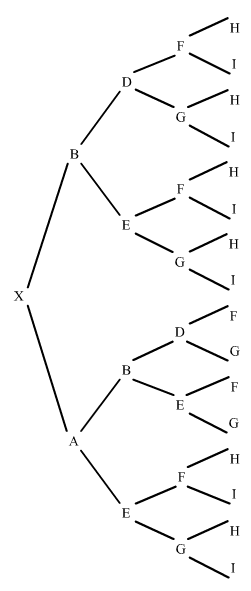 |
|
figura 13
|
Primero busquemos todos los trayectos posibles:
 |
ahora solo falta calcular el trayecto de B, pero como sabemos que es endógamo y su coeficiente de consanguinidad visto en el ejemplo anterior es 0,375:
trayecto de B, desde X hasta A:
Luego sumando los resultados de todos los trayectos nos da:
Fx = 0,5625 = 56,25%, un índice ciertamente muy alto.
La contribución en la endogamia de los antepasados consignados en este ejemplo puede valorarse de la manera siguiente: la mayor contribución pertenece al individuo B; en primer lugar porque sólo se halla a una y dos generaciones de distancia de X, y en segundo lugar por ser endógamo él mismo. La contribución de E también es importante, basicamente por haberse cruzado con su hijo B. Si B es un perro excepcional, X podrá considerarse un buen reproductor a causa de la contribución de B, ya que éste tiene un coeficiente de endogamia bastante alto (37,50).
| ¿ES CONVENIENTE O NO LA CONSANGUINIDAD?. |
La pregunta sobre la conveniencia de la endogamia no es fácil de contestar. Sin embargo, hemos de considerar que si un criador desea fijar unos caracteres y obtener perros con características propias y uniformidad en la descendencia es necesario cierto grado de endogamia. La razón fundamental es que una raza pura o una casta homocigótica no puede obtenerse por otros medios. La selección por sí sola no puede hacerlo. Puede hacerlo atípicamente cuando se ha seleccionado un tipo de padres que presenta fenotipos similares, en cuyo caso, sólo algunos individuos de la progenie tenderán de cuando en cuando a parecerse a los padres.
La endogamia es necesaria para "fijar los caracteres que con tanta perseverancia se han estado seleccionando". Sin duda alguna la endogamia es básica para estabilizar los resultados de la selección.
Pero, ¿cuando se ha de empezar la endogamia?. Principalmente los avances más significativos de la selección se realizan en los primeros estadios. La disminución de los resultados acaba por volverse estacionaria y es más dificil alcanzar nuevos progresos. Entonces, antes de que se diluyan los caracteres seleccionados, es cuando hemos de empezar la endogamia para consolidar los resultados obtenidos. Esta es la filosofía simple y pura de la reproducción: la selección debe suspenderse y la endogamia iniciarse cuando el criador lo considere oportuno.
| SIGNIFICADO DE "LINEA" COMO CONSECUENCIA ENDOGAMICA. |
El término "línea de sangre" muchas veces se usa indiscriminadamente y con cierta frivolidad para indicar un simple parentesco entre los perros. Para algunos, basta para que un reputado semental se aparee con un determinado número de hembras y algunos de sus descendientes sobresalgan en las exposiciones, para hablar de "línea de tal" o "línea de cual". Pero, desde luego, hay algo más.
El término no debe ser usado a la ligera sino que debe reservarse para un criadero con rasgos únicos que se repiten generación tras generación, fruto y culminación de años de cría selectiva ininterrumpida, acompañada de cierta cantidad de endogamia para asegurarse la tendencia hereditaria.
| VENTAJAS Y DESVENTAJAS DE LA ENDOGAMIA. |
Sabemos que existen bastantes perros con cierto grado de endogamia. Pero coeficientes inferiores a 0,25 raramente serán peligrosos. A partir de esta cifra y por encima de 0,5 denota una endogamia de moderada a alta y el perro debe ser vigilado de cerca.
Una de las objeciones con más frecuencia formuladas contra la endogamia es que ésta produce anomalías. La objeción es fundamental y merece ser contestada. En principio hay que precisar que la posibilidad de que se dé una anomalía es muchas veces exagerada. En primer lugar, porque la endogamia por sí misma no puede inducir anomalías; aunque puede suscitar la aparición de cualquier anomalía latente en la perrera.
El apareamiento aleatorio o el cruzamiento externo tienden a mantener a raya los genes detrimentales recesivos salvo apariciones insólitas o inesperadas. Es posible, incluso, que un gen deletéreo sea introducido en la perrera, persista durante algunas generaciones y luego aparezca en una generación sin que nadie entienda qué es lo que está ocurriendo.
Con la endogamia la situación es diferente puesto que tiende a "destapar" los genes recesivos indeseables y por tanto revela las anomalías latentes. En este sentido puede considerarse como una operación purificadora puesto que además de propiciar la aparición de la anomalía, contribuye, igualmente, a su rápida eliminación. Sucede así, porque los individuos en los que aparezcan genes letales en homocigosis no tendrán viabilidad. Y en el caso que afloren genes deletéreos en homocigosis, se evitará su transmisión no haciendo procrear a los individuos portadores de estas anomalías.
No obstante, la aparición aleatoria de una anomalía es infrecuente e imprevisible y no debe confundirse con la "depresión endogámica".
| DEPRESION ENDOGAMICA. |
La llamada "depresión endogámica" es una posibilidad que no puede ignorarse. Con frecuencia se indica que la endogamia produce un decaimiento de la fuerza vital de los reproductores; en efecto, la mayoría de genes recesivos deletéreos se encuentran ocultos en los heterocigotos. Como la consanguinidad disminuye la frecuencia de heterocigotos, tiende a desenmascarar estos genes recesivos detrimentales, en forma de homocigotos.
La pérdida de energía vital se debe a la homocigosidad creciente en los poligenes indeseables que controlan la salud, el vigor y la capacidad reproductiva. El efecto de cada uno de ellos es reducido, y pasaría inadvertido, pero el efecto simultáneo de varios de ellos, se traduce en un animal de aspecto enfermizo sin motivo aparente, de retraso en el crecimiento, menos vivaz que sus hermanos de camada y predisposición a todas las enfermedades que se presenten.
En las poblaciones normales esos poligenes ligeramente deletéreos se presentan como heterocigotos y su presencia pasa inadvertida hasta que son puestos en evidencia por la endogamia.
Para explicar porqué las líneas endogámicas tienen reducido su vigor y fertilidad pensemos primero en una población en la que los cruzamientos se den, habitualmente, entre individuos no emparentados. Si en un individuo aparece por mutación un alelo deletéreo recesivo, transmitirá ese alelo a la mitad de sus descendientes. Como el alelo es recesivo, puede permanecer transmitiéndose y extendiéndose durante generaciones en esa población sin que se note nada en el fenotipo. Si se diera un cruce endogámico y dos miembros de la población, portadores del recesivo, tuvieran descendencia entre ellos, según la herencia mendeliana cada descendiente tendría una probabilidad de 1/4 de ser homocigoto recesivo, mostrando entonces el fenotipo aberrante.
Como las mutaciones deletéreas suelen estar a frecuencia baja en las poblaciones, dos individuos cualesquiera de una población grande, aunque puedan llevar mutaciones deletéreas en heterocigosis, tienen una probabilidad muy baja de ser portadores del mismo alelo recesivo. Por ello la frecuencia de aparición de anomalías genéticas en la descendencia de individuos no emparentados es, proporcionalmente, mucho menor que en la de individuos emparentados.
En las líneas endogámicas en las que se fuerza el cruzamiento entre parientes generación tras generación, irán surgiendo homocigotos recesivos para muchos alelos deletéreos o letales, por lo que cada vez los individuos serán más débiles, menos fértiles y las líneas más difíciles de mantener. Las líneas en las que aparezcan letales en homocigosis se perderán, de manera que tras sucesivas generaciones de endogamia forzada las líneas resultantes no tendrán letales, aunque pueden tener numerosos alelos deletéreos en homocigosis.
Cabe preguntarse porqué los alelos recesivos que aparezcan por mutación han de ser deletéreos en homocigosis. La respuesta es doble, primero porque los alelos mutantes representan una variación frente a una situación previamente adaptada. Pensemos, en este ejemplo: si un impresor comete un error al imprimir una hoja de un libro y cambia al azar una coma, una letra o una palabra, es muy difícil que el texto mejore respecto al manuscrito original, casi siempre empeorará. En segundo lugar porque no importa cuantos alelos recesivos "buenos" haya; en cuanto aparezca un solo homocigoto para un alelo "malo", el individuo será peor que si fuera heterocigoto en ese locus. Basta, por ejemplo, un sólo alelo mutante en homocigosis para producir sordomudez, no importa cuantos genes "buenos" haya para otros caracteres.
Eso no significa que todos los perros sean portadores de esos genes indeseables, ni que la endogamia produzca indefectiblemente una descendencia frágil. Ni mucho menos. La endogamia es inocua en los animales inherentemente sanos y normales, pero merece la pena tomar algunas precauciones, tal como la introducción de al menos un elemento de salud fuertemente ponderado en el análisis de las caracterísicas de los animales.
Los poligenes deletéreos pueden afectar a cualquier rasgo de un perro normal, imperceptiblemente al principio, más seriamente después, si no se han tomado las disposiciones convenientes. Puede producirse un descenso del peso al nacer seguido de crecimiento insatisfactorio, alteración de la capacidad reproductora en la forma de falta de interés por el sexo contrario, falta de deseo de copular o disminución de la frecuencia copulatoria. En ambos sexos la madurez sexual se retrasa. Los períodos de celo de la perra son irregulares, el número de cahorros por camada disminuye y las camadas no alcanzan el peso normal. La depresión endogámica suele presentarse paulatinamente, afectando a ciertos animales y a otros no. Por consiguiente, conviene combatirla haciendo procrear exclusivamente los perros más sanos y vigorosos. Si no se logran los resultados esperados sólo resta el recurso a cruzamientos externos. No obstante, se debe estar muy atento con la integración de material genético distinto, ya que puede comportar la introducción de defectos inesperados.
| EFECTOS DE LA SELECCION Y DE LA ENDOGAMIA. |
Los efectos de la selección y de la endogamia son tan trascendentes en la reproducción animal que juzgamos útil contrastarlos de la forma resumida siguiente:
| SELECCIÓN | ENDOGAMIA |
| perpetuación de alguneos genes | fijación mayoritaria de genes |
| disminución reducida de la heteocigosis | disminución ininterrumpida de la heterocigosis |
| aumento de la similitud fenotípica | aumento de la similitud genotípica |
A primera vista podría parecer que las consecuencias de la selección y la endogamia son complementarias, y que basta con que el criador seleccione los animales apropiados y los haga criar entre sí para que los mejores genes se fijen y se obtengan animales de primera calidad. En realidad resulta muy dificil concentrar todos los genes deseables en unos pocos individuos para que puedan fijarse en la descendencia. La endogamia conlleva el agotamiento de la variabilidad genética y el perro deja de progresar. Una posible solución de compromiso consistirá en una endogamia leve o moderada. En cambio no puede existir compromiso en materia de selección; esta debe ser lo más estricta posible, habida cuenta de la calidad de los animales disponibles. Las fijaciones de los genes deben proseguir incansablemente pero sin prisas, evitando que los rasgos indeseables se fijen antes que el criador haya llegado a eliminarlos. La aparición de ciertos caracteres indeseables son posibles en cualquier momento del proceso, de ahí la necesidad de proseguir la selección.
JOAN FERRER i SIRVENT
Bibliografía: Genética Veterinaria. F.W. Nicholas
Genética, Fundamentos y Perspectivas. M.J. Puertas
Genética para Criadores de Perros. Roy Robinson.
Del Cromosoma al Gen. Institut de Bioquímica Clínica.
|
JOAN FERRER i SIRVENT
LA BORDA D'URTX 17538 URTX (Girona) CATALUNYA Telf: +34-629-613399 e-mail: joanferrer@labordadurtx.org |